
Leslie F. Noé*
Cuenta la leyenda que, en las oscuras aguas del Lago Ness, en Escocia, oculto a la mirada de todos, habita una criatura conocida popularmente como Nessie. Ha sido bautizado Nessieteras rhombopteryx por quienes creen que se trata de un monstruo marino de cuello largo, al que se ha considerado un superviviente prehistórico y cuya anatomía se ha popularizado a lo largo de la historia [1]. La fascinación por esta criatura ha crecido desde que la noticia se reportó por, primera vez, en el periódico escocés el Inverness Courier,c en 1868; consecuentemente, han surgido un sinnúmero de libros, mitos y películas de Hollywood [2].
Sin embargo, aunque la idea suene encantadora, este fantasioso y críptico animal solo es parte del folclore mundial. Sus contrapartes reales, que tienen características muy similares al mítico monstruo del Lago Ness, habitaron los mares del Mesozoico (entre 201 y 65 Millones de años atrás) en Colombia, pero se extinguieron hace 65 millones de años [3]. Estamos hablando de los plesiosaurios, reptiles marinos que medían entre dos y 15 metros de longitud, y presentaban características físicas muy similares a Nessie [4].
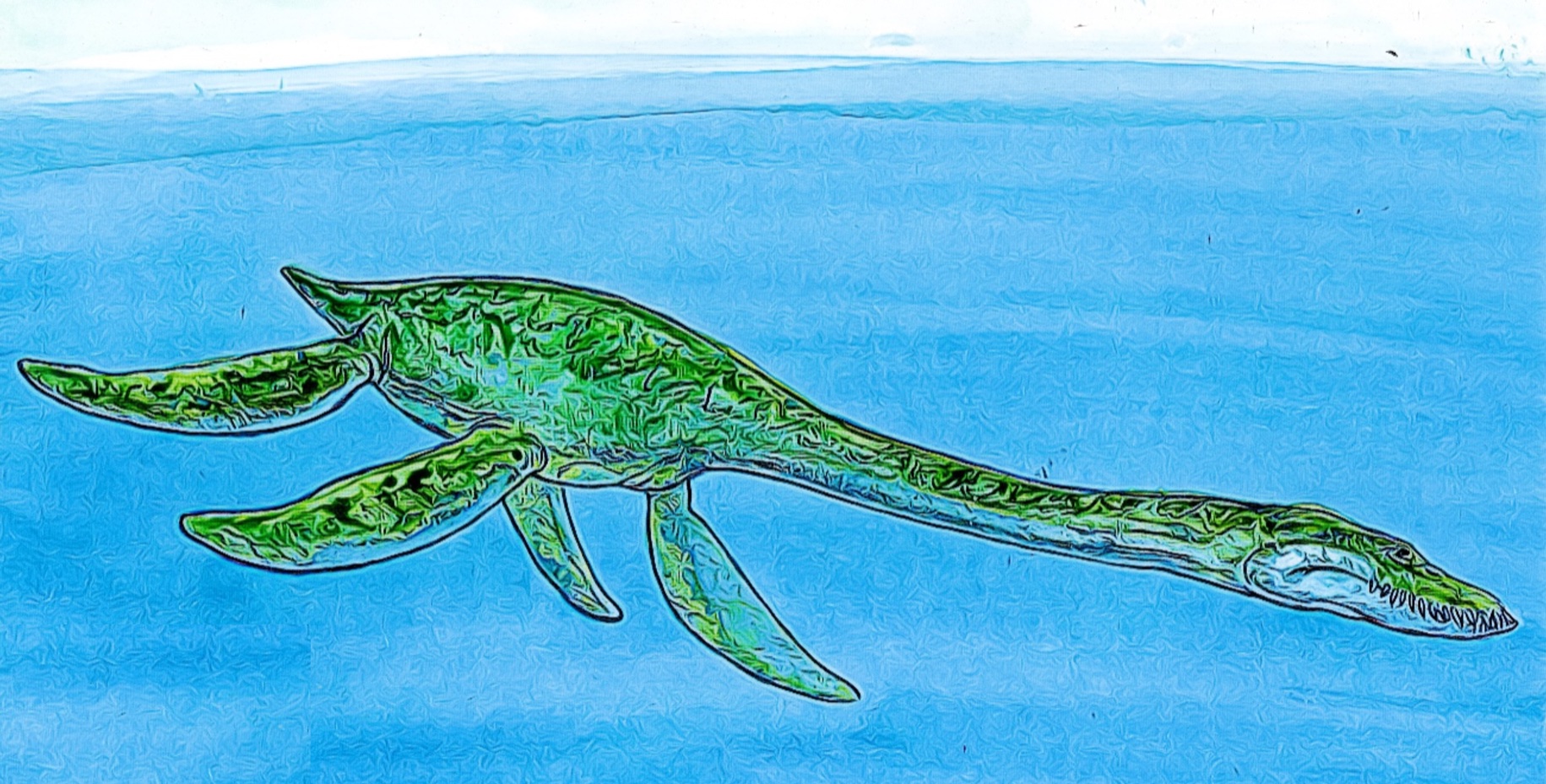
Reconstrucción de un plesiosaurio, la especie colombiana Callawayasaurus colombiensis. Ilustración de Andrés Chaparro, con permiso y copyright © 2021 de Andrés Chaparro y del Museo Geológico Nacional José Royo y Gómez, Servicio Geológico Colombiano, todos los derechos reservados.
Datos curiosos
Existe mucha controversia con respecto a la existencia de un verdadero monstruo del Lago Ness. Los turistas acuden a este legendario lugar en Escocia esperando avistar el monstro, resultando en una gran cantidad de dudosa evidencia respaldando la existencia de este gran monstruo del Mesozoico. Lo cierto es que, a pesar de la supuesta evidencia fotográfica, el registro fósil existente evidencia que los plesiosaurios no podían doblar su cuello arriba como un cisne, lo cual es una parte fundamental de la leyenda del monstruo del lago Ness [5].
El primer registro científico reportado de un plesiosaurio ocurrió, en 1821, a cargo de Mary Anning, en el sur de Inglaterra. Desde entonces, numerosos ejemplares han sido encontrados mundialmente, algunos de ellos muy bien preservados. Gracias a su exitosa supervivencia durante el Mesozoico, hay registro de fósiles de plesiosaurios en todos los continentes alrededor del mundo, incluyendo Colombia.
La alta cantidad de fósiles encontrados ha permitido realizar análisis de variabilidad dentro de las especies, haciendo posible la comparación entre especímenes determinados [6]. Por ejemplo, el fósil de plesiosaurio más grande registrado, que se encuentra en el libro de los Guinness World Records, es atribuido a un Liopleurodon, hallado en el Reino Unido, que puedo haber alcanzó una longitud de 25 metros.
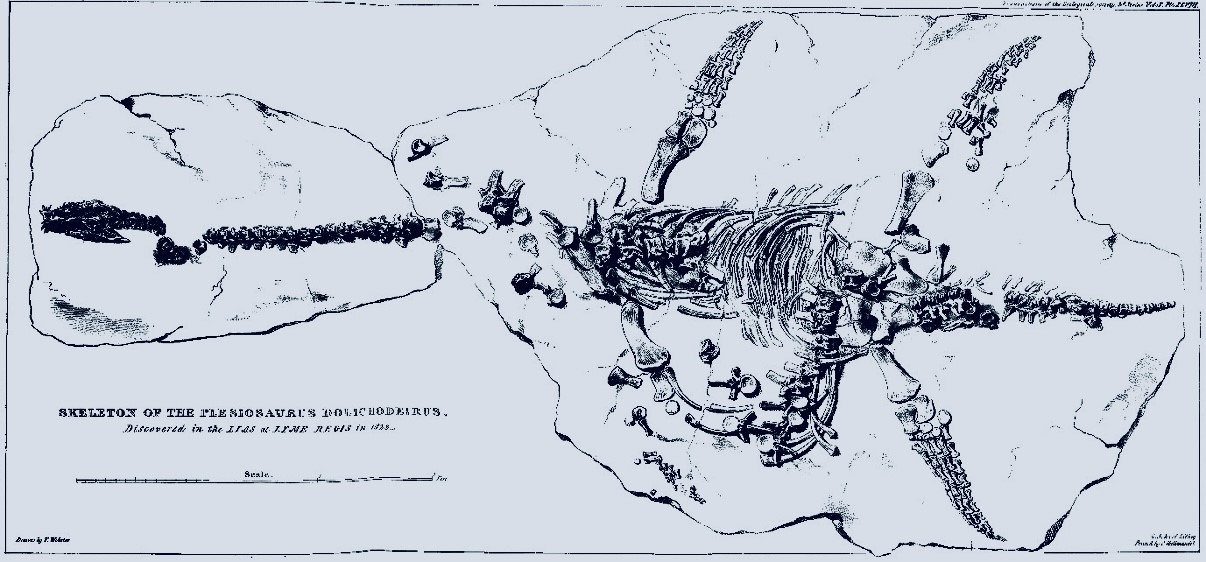 El primer esqueleto completo de un plesiosaurio, descubierto en Lyme Regis, en la costa sur de Inglaterra. Descubierto por Mary Anning y publicado por William Conybeare en 1821.
El primer esqueleto completo de un plesiosaurio, descubierto en Lyme Regis, en la costa sur de Inglaterra. Descubierto por Mary Anning y publicado por William Conybeare en 1821.
¿Cómo se movían estos gigantes?
Los plesiosaurios fueron únicos entre los vertebrados que utilizaron dos pares de aletas (normalmente con las delanteras un poco más largas que las traseras) para moverse a través de los océanos del Mesozoico [7]. A partir de los restos fósiles y modelos computacionales, se ha determinado que el modo de natación de estos reptiles marinos se parecería al de algunos tetrápodos (que tienen cuatro patas) que 'vuelan' bajo el agua, como las tortugas marinas y los pingüinos [6]. Adicionalmente, se piensa que las extremidades únicas de los plesiosaurios podían usar los flujos oceánicos turbulentos a su favor, y que las extremidades anteriores y posteriores les permitirían moverse velozmente y con gran control cuando buscaban comida [9].
Se cree que las puntas de las extremidades se movían en forma de ocho, generando empuje durante todo el ciclo de movimiento, como las tortugas o los leones marinos en la actualidad. Sin embargo, las aletas anteriores y posteriores de los plesiosaurios podrían haber trabajado ligeramente fuera de fase [8]. Permitiéndoles seguir una trayectoria oblicua para reducir la fricción dentro del agua y poder sumergirse o subir a la superficie fácilmente, ganando velocidad, potencia y ligereza al desplazarse [9].
Sabemos que los plesiosaurios podían mover su largo cuello hacia abajo con mayor facilidad que en cualquier otra dirección. Las costillas a ambos lados del centrum (el cuerpo central) de las vértebras dificultarían el movimiento lateral, y la espina neural en la parte superior del centrum restringiría severamente el movimiento hacia arriba. Además, el cuello tampoco era lo suficientemente flexible para realizar movimientos bruscos, necesarios (por ejemplo) para la cacería, ya que sus uniones eran esencialmente planas [5, 10].
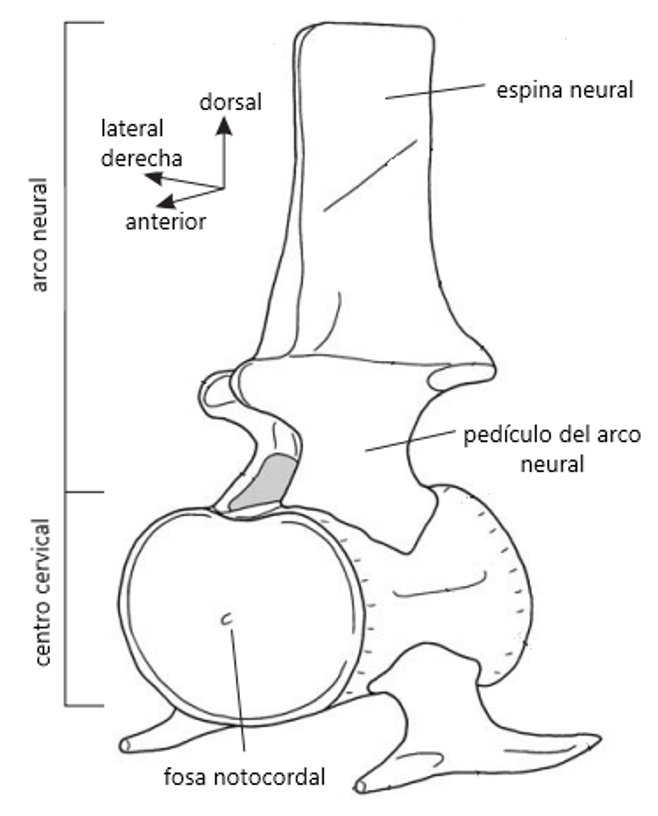
Esquema de una vértebra cervical (del cuello) de un plesiosaurio. Modificado de [5], dibujado por Dr. L.F. Noè.
Durante mucho tiempo, el cuello largo del plesiosaurio se consideró muy útil para la respiración por fuera del agua, como si se tratara de un snorkel. Así pues, el plesiosaurio mantendría su cuerpo sumergido mientras obtenía oxígeno de la atmosfera. No obstante, esta hipótesis ha sido descartada, debido a las diferencias de presión que habría entre el aire en los pulmones y la atmósfera, imposibilitando la respiración del animal. Además, nadar cerca de la superficie implicaría un gran gasto energético al ser un movimiento en contra de las olas [5]. Sin embargo, el cuello largo parece no ser muy práctico para la caza, pero el cuerpo proporciona una plataforma de caza desde la cual el cuello puede alcanzar la comida debajo del cuerpo, posiblemente incluso alimentándose de organismos en el lecho marino [9].
Correspondiente con el largo cuello, la tráquea sería muy extensa, resultando en una mayor acumulación de aire que no participa en el intercambio gaseoso. Esto tendría efectos en el tamaño de los pulmones, que se harían más grandes, o la inhalación-exhalación debería ser más profunda y frecuente, para evitar los problemas asociados a un gran espacio muerto [5]. La tráquea también habría sido angosta, lo que relentizaría la frecuencia respiratoria, generando una contradicción entre la cantidad de oxígeno requerida por el animal y la necesidad de reducir el tiempo dedicado a respirar en la superficie [11].
Plesiosaurios como depredadores
Los plesiosaurios fueron depredadores muy exitosos en los océanos del Mesozoico, lo que ha llevado a diferentes suposiciones relacionadas con su posible comportamiento alimenticio. Sus dientes eran alargarlos, cónicos, curvados y puntiagudos, y las mandíbulas podrían haber funcionado a modo de cuchara [9]. En general, hay evidencia de que los plesiosaurios cazaban peces y cefalópodos de cuerpo blando (similares a la sepia de hoy) [12], encontrándose restos de los mismos en los estómagos de fósiles. Su dieta habría consistido principalmente de animales bentónicos (es decir del fondo del mar), especialmente crustáceos y otros tipos de presas invertebradas [13, 14].
En efecto, los dientes sobresalían periféricamente y entrecruzándose entre sí [15], permitiendo atrapar a la presa detrás de los dientes, como se observa, por ejemplo, en la foca cangrejera (Lobodon carcinophagus). Por eso, se ha determinado que algunos plesiosaurios se alimentaban por tamizado, siendo capaces de procesar grandes volúmenes de agua y de sustratos [16]. De hecho, sus dientes eran largos y puntiagudos, y por lo tanto relativamente débiles, pero espaciados para permitir que el agua y el sustrato fueran expulsados, mientras que las presas eran atrapadas. Por ejemplo, en un fósil del plesiosaurio Morturneria se encontró que había probablemente una válvula capaz de sellar la narina interna, lo que permitiría que algunos plesiosaurios pudieran ingerir el sustrato sin que el agua o los sedimentos entraran a las fosas nasales [16].
La morfología y la extensión del cuello de los plesiosaurios está dada por la relación entre la longitud y el grosor del mismo, siendo mucho más largo que ancho [4]. Esto permite una estimación del tamaño promedio que podría tener una presa para ser engullida sin que se presentara asfixia. Basado en esto, puede decirse que los plesiosaurios se alimentarían de organismos relativamente pequeños, lo cual ocurriría muy frecuentemente para poder suplir sus necesidades energéticas. Esto contrasta con sus parientes cercanos, los pliosaurios cabezones, que podrían haberse alimentado de animales como cefalópodos, belemnites, amonitas y otros organismos de mayor tamaño, como peces grandes y reptiles [14].
El cuello de los plesiosaurios probablemente condicionaba su forma de nado, por lo que el organismo pudo haber desarrollado estrategias de caza acordes con su morfología. Se proponen algunos modelos con lo que se intenta explicar el éxito de los plesiosaurios como depredadores activos. Los primeros estudios realizados sobre la movilidad del cuello aparentemente han mostrado que estos presentaban un gran rango de movimiento lateral, lo que les daría un mayor campo de acción o reacción al movimiento de sus presas. Más recientemente, se determinó que el rango de movimiento del cuello estaba restringido más a la dirección ventral que a hacia la dorsal o lateral, es decir, los plesiosaurios podían mover su cuello más fácilmente hacia abajo que hacia arriba o por los lados [5, 17].
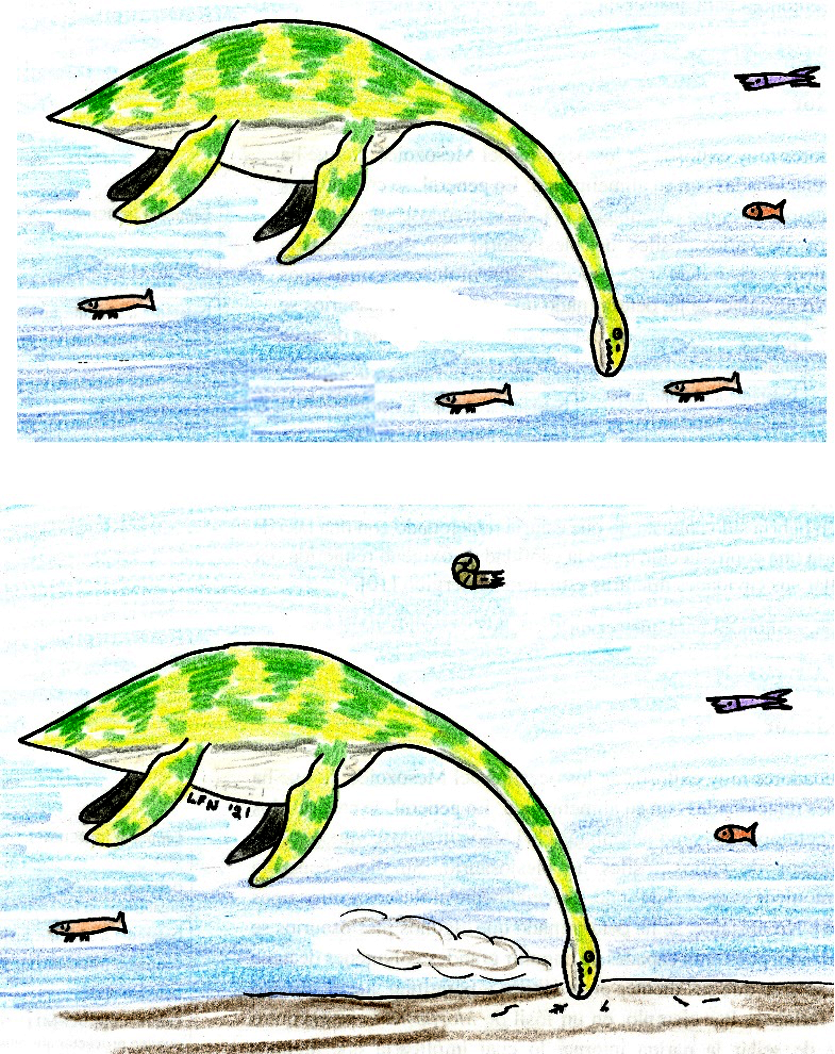
En algunos fósiles de elasmosaurios (un tipo de plesiosaurio con cuello extremadamente largo) se han encontrado muchos gastrolitos. Gastrolitos (o piedras estomacales) son rocas redondeadas contenidas dentro de la vía digestiva de un animal, que contribuyen a la trituración de los alimentos. Aunque inicialmente se creía que estaban relacionados únicamente con la flotabilidad del animal (como en los cocodrilos modernos), estudios recientes encontraron que la relación entre el peso total de los gastrolitos y la masa corporal del animal no influye significativamente en la flotabilidad de los plesiosaurios [18].
Se ha propuesto, por lo tanto, que los gastrolitos ayudaban en la digestión, actuando como pseudo-dientes en un estómago musculoso. Además, los dientes de los vertebrados marinos generalmente están adaptados para atrapar, pero no masticar, a sus presas, por lo que puede determinarse que los plesiosaurios usaban los gastrolitos para suplir esta función. Adicionalmente, los gastrolitos podrían ser una fuente de minerales, y la descomposición de los mismos dentro del cuerpo liberaría nutrientes esenciales que los animales no podían adquirir a través de la dieta [19].
A pesar de que la flotabilidad no habría sido la función principal, los gastrolitos habrían contribuido, al igual lo hacen en los cocodrilos modernos, ayudando al animal a permanecer parcialmente sumergido [20]. Adicionalmente, en los plesiosaurios los gastrolitos podrían haber conferido la capacidad de maniobra con mayor precisión cuando se movían lentamente por el medio acuático [20].
En resumen, podemos inferir que los plesiosaurios fueron cazadores exitosos dentro su medio ambiente, y que algunos se alimentaban en la región bentónica del lecho marino y usaban gastrolitos para ayuda con la digestión de sus presas, y controlar su flotabilidad y aumentar la capacidad de maniobra.

Kimmeridgia Antiquior (Una Kimmeridge Más Antigua) que representa la vida en los mares del Jurásico Tardío del sur de Inglaterra. Anota los plesiosaurios de cuello largo como depredadores y presas. Publicado por primera vez en [21]. Ilustración por Robert Nicholl, copyright © 2016 Universidad de los Andes, Museum of Jurassic Marine Life, Leslie Noè y Robert Nicholls, todos los derechos reservados.
Nacimiento, crecimiento y enfermedades
La evidencia fósil ha mostrado que los plesiosaurios eran organismos vivíparos, o animales cuyo embriones se desarrollan dentro el vientre de la hembra, en lugar de nacer de un huevo. Los plesiosaurios presentaban un estrategia K de reproducción, produciendo un bajo número de crías con una alta inversión parental en cada bebé. Esto es relativamente extraño en los reptiles, que tienden a presentar con selección-R, con un alto número de crías y sin cuidado de los padres. La estrategia K también se ha encontrado en otros reptiles marinos, como los ictiosaurios (reptiles marinos mesozoicos en la forma de un pez) que gestan hasta ocho crías al tiempo [22].
En ictiosaurios, las crías nacían con la cola primero, como en delfines y ballenas día de hoy, reduciendo así la posibilidad que la cría se ahogue durante el parto. Este también podría haber sido el caso de los plesiosaurios, aunque en este caso producían un numero menor de crías de tamaño mayor.
La viviparidad de los plesiosaurios, junto con otras adaptaciones a la vida marina, como las grandes extremidades hidrodinámicas y su largo cuello, permiten determinar que estos organismos no necesitaban desplazarse a tierra para construir nidos y poner huevos en ellos. Un huevo necesita ser puesto en tierra para el desarrollo del embrión, ya que el intercambio gaseoso asociado a la respiración del mismo es imposible en el agua [24].
A diferencia de los humanos, y la mayoría de los mamíferos que tienen solo siete vértebras cervicales, los plesiosaurios se desarrollaron agregando nuevas vértebras con algunas especies presentando hasta 70 vértebras, haciendo la zona cervical mucho más larga que el cuerpo [25]. Los plesiosaurios se caracterizaban por tener tres etapas asociadas al desarrollo osteológico de sus vértebras: i) la etapa juvenil, que se distinguía porque los arcos neurales de las vértebras no están fusionados al centrum (equivalente a nuestros años de preadolescencia); ii) la etapa adulta, identificable porque los arcos neurales de las vértebras se han fusionado pero la unión es visible (equivalente a nuestros años reproductivos); y iii) la etapa senil, en la cual hay evidencias de osificación avanzada y de la fusión de los arcos neurales y el centrum sin línea de conexión visible (equivalente a nuestra vejez), a menudo con evidencia de enfermedades óseas degenerativas [26].
Los plesiosaurios muestran evidencia de tasas de crecimiento óseo muy rápidas. El rápido crecimiento pre-reproductivo a menudo se asocia tanto con el cuidado parental de los jóvenes como con los altos niveles metabólicos de los organismos. Esto implica un elevado gasto energético y, por lo tanto, requiere de la adquisición continua y activa de presas, y un metabolismo endotérmico. La endotermia se asocia normalmente con los mamíferos, más que con los reptiles, y es la capacidad de un organismo de mantener su temperatura corporal independientemente del medio ambiente en el cual se encuentren. Esto es importante, ya que el agua tiene una alta capacidad calorífica, por lo que disipa rápidamente el calor de un cuerpo (esta es la razón por la que una piscina se siente fría). El desarrollo de la viviparidad, como modo de reproducción, se asocia actualmente a reptiles que viven en ambientes estacionalmente fríos, como las serpientes [27]. Muchas adaptaciones fisiológicas de los plesiosaurios parecen indicar un regreso al agua de reptiles terrestres.
La patología (o paleopatología, el estudio científico de enfermedades antiguas) de los plesiosaurios es considerada como una gran fuente para entender el comportamiento y la forma de vida. Hay estudios que reportan que los plesiosaurios sufrieron del síndrome de descompresión (o Enfermedad de Caisson), en el que los cambios bruscos de presión hace que se formen burbujas de nitrógeno en la sangre. Estas burbujas de nitrógeno pueden formarse o migrar a muchos tejidos, pero se pueden observar en el registro fósil como necrosis avascular; es decir, la muerte del hueso, por la pérdida del suministro de sangre, con su consecuente fractura y colapso. Los extremos de los huesos largos son especialmente susceptibles, y el hecho de que algunos plesiosaurios padecieran de Enfermedad de Caisson indica que cambiaban de forma abrupta la profundidad en la que buceaban, ya sea por la búsqueda de presas o por huir de depredadores [28]. Sin embargo, estudios morfológicos indican que esta enfermedad es rara en el registro fósil e indicando que la mayoría de plesiosaurios habría vivido en hábitats costeros de aguas poco profundas [28].
¿Dónde se encuentran los plesiosaurios más importantes en Colombia?
Una gran parte de los reptiles marinos hallados en Colombia se han encontrado en la región de Ricaurte Alto, alrededor de Sáchica, Sutamarchán y Villa de Leyva, en Boyacá. A partir de reconstrucciones paleogeográficas, se ha establecido que el área alrededor de Villa de Leyva era un mar somero, de hasta 130 metros de profundidad, en el Cretácico. Durante millones de años esta área fue hábitat de reptiles marinos, incluyendo plesiosaurios, ictiosaurios, tortugas y muchos otros organismos marinos.

Callawayasaurus colombiensis, el esqueleto más completo de un plesiosaurio descubierto hasta ahora en Colombia. Se trata de un animal de aproximadamente 8 m de largo descubierto en la región Ricaurte Alto de Boyacá. Imagen con permiso y copyright © 2021 del Museo Geológico Nacional José Royo y Gómez, Servicio Geológico Colombiano, todos los derechos reservados.
El Callawayasaurus colombiensis (originalmente nombrado Alzadasaurus colombiensis) es el plesiosaurio más importante encontrado en Colombia. Pertenece a la familia Elasmosauridae, cuyas especies se pueden diferenciar mediante la morfología de su cráneo, basándose en la distribución de los huesos, el tamaño de los dientes, sus posiciones en las mandíbulas, y por el tamaño del cuello [29].
Callawayasaurus tiene un cráneo de 35 cm de largo y un cuerpo de 8 m de longitud, y hay un espécimen en exhibición en el Museo Geológico Nacional José Royo y Gómez, del Servicio Geológico Colombiano, en Bogotá. El cráneo de este fósil se prestó a científicos de la Universidad de Berkeley en dos ocasiones para realizar varios estudios. Sin embargo, no fue devuelto, por ello, se dejó en exhibición una réplica hecha en resina. En 2018 el cráneo fue solicitado por la paleontóloga Marcela Gómez, quien, junto con el profesor Leslie Noè en Uniandes, hicieron las investigaciones y procesos necesarios para su repatriación. Ahora el cráneo se puede visitar en la vitrina contigua al esqueleto.

El cráneo de Callawayasaurus colombiensis regresó del Museo de Paleontología de la Universidad de California (Berkeley, EE. UU.) luego de un préstamo de más de 60 años. El cráneo se exhibe ahora en el Museo Geológico Nacional José Royo y Gómez. Imagen con permiso y copyright © 2021 del Museo Geológico Nacional José Royo y Gómez, Servicio Geológico Colombiano, todos los derechos reservados.
Puede que el monstruo del Lago Ness sea solo una leyenda; sin embargo, durante el Mesozoico, los mares, incluidos los situados en Colombia, estaban habitados por diversas especies de plesiosaurios cuya morfología se ha utilizado como base para la criatura legendaria. Se espera que futuros estudios morfológicos y filogenéticos nos permitan entender más sobre la vida y desarrollo de estos enigmáticos reptiles, además de los procesos geológicos y ambientales que llevaron a su crecimiento poblacional y posterior extinción a nivel regional (para el caso de los especímenes colombianos), y mundial hace 65 millones de años.
Los plesiosaurios son un grupo de reptiles marinos difícil de estudiar, puesto que no existen organismos vivos con los que se puedan comparar ni en morfología, comportamiento o hábitat. Sin embargo el estudio de fósiles en Colombia, y en otros países, permite entender su locomoción, hábitos predatorios, dieta, respiración y reproducción. Estos fascinantes, y ahora extintos reptiles, con sus dos pares de aletas que permiten un rápido nado, y sus enigmáticos cuellos largos habitaban los mares someros alrededor de Villa de Leyva, muy lejos de las gélidas aguas de un lejano Loch escocés.
[1] Sabadell M, El monstruo del lago Ness Muy Interesante, 2019.
[2] Monstruo del lago Ness: el estudio que asegura haber dado con una explicación para el legendario misterio BBC News Mundo, 2019.
[3] Morrone JJ and Fortino AD, La Zoología de los Animales Fantásticos: Apuntes para un Bestiario Criptozoológico Revista de la Museo de Univerisidad la Plata 1996; 8: 75-80.
[4] Benton MJ, The great Sea Dragons. In: Benton MJ Vertebrate Palaeontology, 3rd ed. Blackwell Publishing Ltd 2005.
[5] Noè LF, Taylor MA, and Gómez-Pérez M. An integrated approach to understanding the role of the long neck in plesiosaurs. Acta Palaentologica Polonica 2017; 62(1):. 137–162.
[6] Tutin SL and Butler RJ. Europe PMC Funders Group The completeness of the fossil record of plesiosaurs, marine reptiles from the Mesozoic. Acta Palaentologica Pol 2017; 62(3): 563–573.
[7] Motani R. The Evolution of Marine Reptiles. Evolution and Education Outreach 2009; 2: 224–235.
[8] Carpenter K, Sanders F, Reed B, and Reed J. Plesiosaur Swimming as Interpreted from Skeletal Analysis and Experimental Results. Transactions of the Kansas Academy of Sciences 2010; 113 (1/2): 1–34.
[9] Halstead LB. Plesiosaur locomotion. Journal of the Geological Society, London 1989; 146: 37–40.
[10] Storrs G. Function and phylogeny in Sauropterygian (Diapsida) evolution. American Journal of Science 1993;. 293(A): 63–90.
[11] Buchy M-G, Frey E, and Salisbury SW. The internal cranial anatomy of the Plesiosauria (Reptilia , Sauropterygia): evidence for a functional secondary palate. Lethaia 2006; . 39(1981): 289–303.
[12] McHeny CR, Cook AG, and Wroe S, Bottom-Feeding Plesiosaurs. Science 2005; 310: 75.
[13] Massare JA, Swimming capabilities of Mesozoic marine reptiles: Implications for method of predation. Paleobiology 1988; 2(2): 187–205.
[14] Massare JA. Tooth morphology and prey preference of mesozoic marine reptiles Journal of Vertebrate Paleontology 1987; 7(2): 121–137.
[15] Cabrera Á. Un plesiosaurio nuevo del Cretáceo del Chubut. Revista del Musueo de la Plata 1942; 8: 113–130.
[16] F.R. O’Keefe et al., Cranial anatomy of Morturneria seymourensis from Antarctica , and the evolution of filter feeding in plesiosaurs of the Austral Late Cretaceous Journal of Vertebrate Paleontology, vol. 4634, no. 37, p. 4, 2017.
[17] Nagesan RS, Henderson DM, Anderson JS. A method for deducing neck mobility in plesiosaurs, using the exceptionally preserved Nichollssaura borealis. Royal Society Open Science 2018; 5: 172307.
[18] Cerda IA, Salgado L. Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Patagonia, Argentina. Resvista la Asociacion Paleontológica Argentina 2008; 45(3): 529–536.
[19] O’Keefe FR and Carrano MT. Correlated trends in the evolution of the plesiosaur locomotor system Paleobiology 2005; 31(4): 656–675.
[20] Rothschild B. Decompression syndrome in plesiosaurs (Sauropterygia: Reptilia). Journal of Vertebrate Paleontology 2003; 23(2): 324–328.
[21] Noè LF, Gómez-Pérez M, Nicholls R. Mary Anning, Alfred Nicholson Leeds and Steve Etches. Comparing the three most important UK ‘amateur’ fossil collectors and their collections. Proceedings of the Geologists' Association 2019; 130(3-4): 366-389
[22] Motani R, Jiang DY, Tintori A, Rieppel O, Chen GB. Terrestrial origin of viviparity in Mesozoic marine reptiles indicated by early triassic embryonic fossils. PLoS One 2014, 9(2): e88640
[23] O’Keefe FR, Chiappe LM. Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia) Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia). Science 2011; 333: 870–873.
[24] Liu J, Organ CL, Benton MJ, Brandley MC, Aitchison JC. Live birth in an archosauromorph reptile. Nature Communications 2017; 8: 1–8.
[25] Wintrich T, Hayashi S, Houssaye A, Nakajima Y, Sander PM. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances 2017; 3(12): 1–11.
[26] Garat LM. Análisis comparativo de la microestructura ósea y paleohistología en diferentes secciones de la columna vertebral de un plesiosaurio (elasmosáurido) del Cretácico Superior de Antártida. Univesidad Nacional de Río Negro 2019.
[27] Fleischle CV, Wintrich T, Sander PM. Quantitative histological models suggest endothermy in plesiosaurs. PeerJ 2018; 6: e4955.
[28] Rothschild B, Clark N, Clark C. Evidence for survival in a Middle Jurassic plesiosaur with a humeral pathology: What can we infer of plesiosaur behaviour?. Palaeontologia Electronica 2018; 21(16): 1-11.
[29] Carpenter K. Revision of North American Elasmosaurs from the Cretaceous of the Western Interior. Paludicola 1999; 2(2): 148–171.
* Departamento de Geociencias, Universidad de los Andes